Tendinopathies are painful, disabling conditions that afflict 25% of the adult human population. Filling an unmet need for realistic large-animal models, we here present an ovine model of tendon injury for the comparative study of adult scarring repair and fetal regeneration. Complete regeneration of the fetal tendon within 28 days is demonstrated, while adult tendon defects remained macroscopically and histologically evident five months post-injury. In addition to a comprehensive histological assessment, proteome analyses of secretomes were performed. Confirming histological data, a specific and pronounced inflammation accompanied by activation of neutrophils in adult tendon defects was observed, corroborated by the significant up-regulation of pro-inflammatory factors, neutrophil attracting chemokines, the release of potentially tissue-damaging antimicrobial and extracellular matrix-degrading enzymes, and a response to oxidative stress. In contrast, secreted proteins of injured fetal tendons included proteins initiating the resolution of inflammation or promoting functional extracellular matrix production. These results demonstrate the power and relevance of our novel ovine fetal tendon regeneration model, which thus promises to accelerate research in the field. First insights from the model already support our molecular understanding of successful fetal tendon healing processes and may guide improved therapeutic strategies.
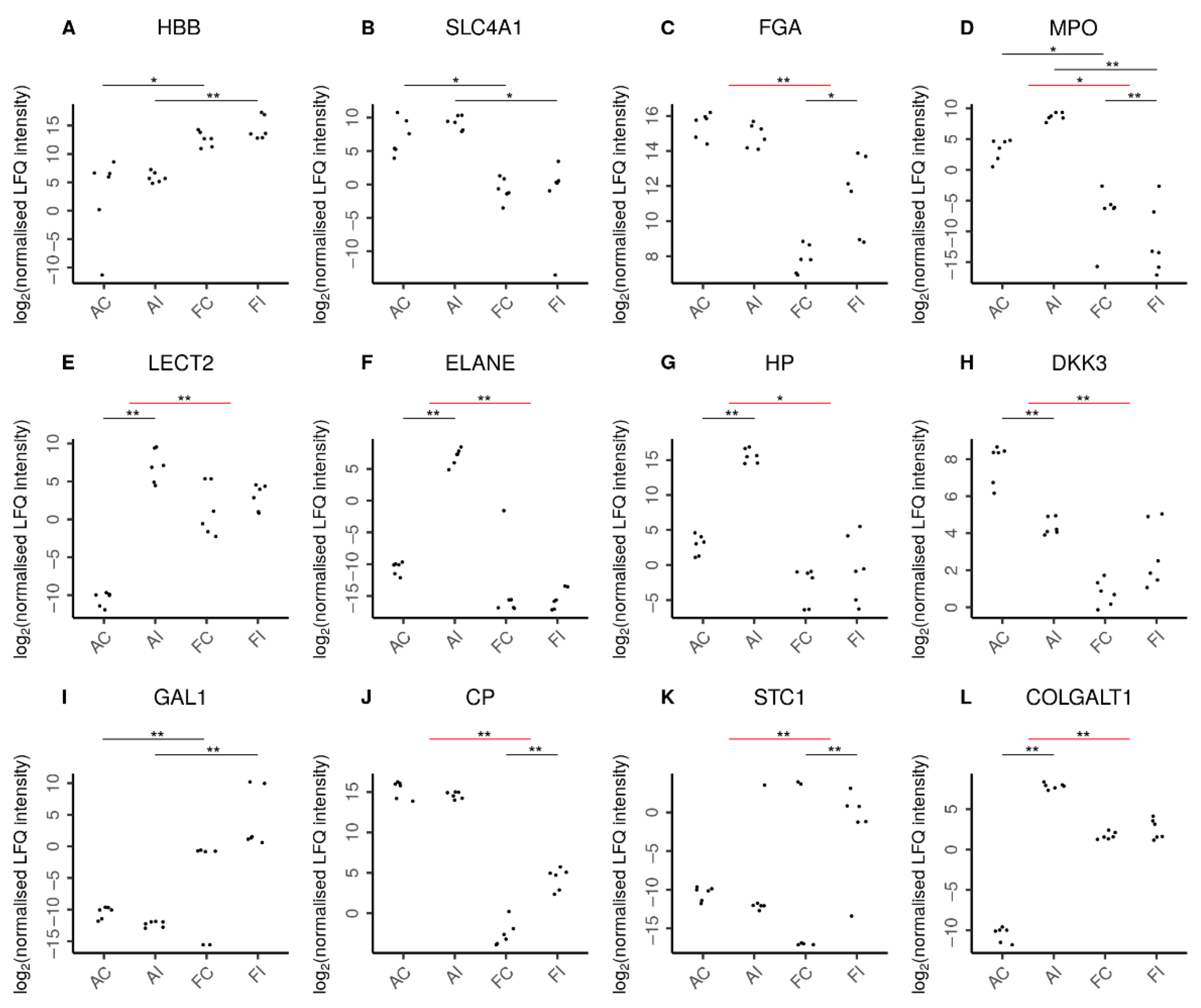
Figure 7. Selected proteins indicating characteristic features in adult and fetal animal samples. Log2 normalized LFQ intensities of each protein are shown for adult control (AC), adult injured (AI), fetal control (FC), and fetal injured (FI) tendon samples. The pairwise comparisons discussed are indicated by horizontal bars. Comparisons can be between two samples directly (black bars) or between the adult and fetal responses to injury (red bars). Statistical significance is then shown by double asterisks (**) and marginal statistical significance by a single asterisk (*). In line with expectations, (A) higher amounts of hemoglobin fetal subunit beta (HBB) were seen in fetal samples. Similarly, (B) higher levels of solute carrier family 4 member 1 (SLC4A1) reflected the increased presence of erythrocytes in adult samples. Remarkably, (C) fibrinogen chain alpha (FGA) was specifically increased only in fetal injury. Adults featured a higher presence of neutrophils (D) as reflected by myeloperoxidase (MPO) levels in these tissues. Moreover, the adult-specific response to injury was characterized by chemoattraction of neutrophils (E) as indicated by leukocyte cell-derived chemotaxin 2 (LECT2) and the secretion of antimicrobial proteins (F) by neutrophils such as neutrophil elastase (ELANE). In addition, the adult-specific response to injury featured up-regulation of pro-inflammatory proteins (G) such as the acute phase haptoglobin (HP) as well as the down-regulation of the anti-inflammatory (H) dickkopf WNT signaling pathway inhibitor-3 (DKK3). In contrast, fetal samples featured higher levels of anti-inflammatory proteins (I), such as galectin-1 (GAL1). Moreover, the fetal-specific response to injury featured up-regulation of anti-inflammatory proteins such as (J) the inflammation-resolving ceruloplasmin (CP) and (K) stanniocalcin-1 (STC1). The regulation of extracellular matrix proteins also played a more pronounced role in the adult response to injury with over a dozen ECM proteins specifically up-regulated in adults, as exemplified by (L) the collagen beta(1-O)galactosyltransferase 1 (COLGALT1). Supplementary Figure S1 shows further proteins characterizing the adult and fetal responses to injury.